nach Dr. med. Heinrich Kremer

Cellsymbiosis Therapie:
nach Dr. Kremers Postulat: Paradigmenwechsel in der Behandlung chronischer Leiden
Die Bezeichnung „Cellsymbiosis“ (Cellsymbiosistherapie) leitet Dr. med. Kremer von der Vorstellung ab, dass sich die menschliche Zelle im Laufe der Evolution aus zwei gegensätzlichen Zellen durch eine Zell-Fusion zu einer Zell-Symbiose entwickelt habe. Die gemeinten Zellen sind einerseits Archaea-Bakterien mit sowohl anaeroben und aeroben (ohne und mit Sauerstoff) Stoffwechsel, die das Zellplasma bilden sollen und Proteo- Bakterien, die aerob die Energiegewinnung bewerkstelligen und als Endobionten die späteren Mitochondrien gebildet haben sollen.
Nach der Kremerschen Lehre der Cellsymbiosis hat die Entstehung von chronischen Erkrankungen größtenteils ihre Ursache in einer gestörten Mitochondrienfunktion, die jedoch nicht mit den Mitochondriopathien in der wissenschaftlichen Medizin verwechselt werden dürfen. Nach dieser Lehre sind die häufigsten Zivilisationskrankheiten wie Herzinfarkt, Schlaganfall, Arteriosklerose, Hypertonie, Diabetes mellitus usw einzig die Folge von Mitochondrienfunktionsstörungen. Ziel einer „Cellsymbiosistherapie“ soll es sein, eine Verbesserung oder Wiederherstellung der Mitochondrienfunktion zu erreichen.
Der Ausweg aus der therapeutischen Sackgasse
Artikel von Dr. Kremer
Im Juli 2003 „stürzte der genetische Himmel ein“, wie ein Forscher resümierte. Was war passiert? Auf dem internationalen Kongress für Genetik in Melbourne hatten Genforscher aus aller Welt „das Ende des Anfangs der Genomforschung“ ausgerufen. Zuvor waren die endgültigen Ergebnisse eines der ehrgeizigsten Forschungsprojekte in der modernen Medizin publiziert worden. Seit Ende der 80er Jahre hatten internationale Forschungsgruppen im Verbund sämtliche Gene in dem mehr als 3 Milliarden Einzelbausteine umfassenden Riesenmolekül der DNA im menschlichen Zellkern katalogisiert. Eingesetzt wurden computergestützte, automatisierte Sequenziermaschinen. Die Erwartung war, dass im menschlichen Genom mindestens 120000 Gene vorhanden sein müssten, also besondere Abschnitte in der DNA mit einer verschlüsselten Reihenfolge der DNA Bausteine, den 4 klassischen sog. Nukleobasen Adenin (A), Guanosin (G), Cytosin (C) und Thymin (T). Die Annahme basierte auf der Tatsache, dass es in den menschlichen Zellen mehr als 100000 Proteine gibt, die für ihre Synthese außerhalb des Zellkerns eine genetische Blaupause benötigen. Hinzurechnen musste man etwa 20.000 regulierende Gene, die erforderlich sind, um den gesamten Prozess der Abschrift der Gene in eine mobile RNA-Synthesevorlage bis zum fertigen Protein, die sog. genetische Expression, zu steuern. In einem Parallelprogramm sequenzierten Genforscher die Gene im DNA-Molekül in den Zellkernen von Mauszellen.
Die Ergebnisse waren schockierend: Das menschliche Zellkerngenom besitzt etwas mehr als 25.000 Gene, das der Maus 24.000. Inzwischen sprechen Genforscher von nur noch 21.000 humanen Zellkerngenen. Das sind kaum mehr Zellkerngene als in einem der Haustierchen der Genforschung, einem winzigen Fadenwurm von wenigen Millimeter Länge und exakt 969 Zellen, gefunden wurden. Im Vergleich dazu besitzt der Mensch geschätzte 50 Billionen Zellen. Verhältnismäßig einfache Pflanzen, wie die Ackerschmalwand, weisen dagegen ein Mehrfaches an Zellkerngenen auf als die menschlichen Zellkerne.
Der Nobelpreisträger David Baltimore, einer der bis dahin weltweit anerkanntesten Meinungsführer des genetischen Determinismus der menschlichen Existenz, hatte in einem geradezu verzweifelten Kommentar zu den 2001 publizierten vorläufigen Ergebnissen des Humanen Genomprojektes festgestellt: „Falls im menschlichen Genom nicht noch viele Gene vorhanden sind, die unsere Computer nicht erkennen können, müssen wir zugeben, dass wir unsere im Vergleich zu Würmern und Pflanzen zweifellos größere Komplexität nicht durch ein Mehr an Genen gewonnen haben. Die Erkenntnis dessen, was uns unsere Komplexität verleiht, … bleibt eine große Herausforderung für die künftige Forschung.“ (Baltimore, D. (2001), Our genome unveiled. Nature 409:814 16)
 Was Baltimore und die große Mehrheit seiner Kollegenschaft nach dem Zusammenbruch des genetischen Weltbildes nicht sagen, ist die fundamentale Tatsache, dass alle grundlegenden Theorien der gentechnisch fixierten modernen Medizin zur Zellenergie, Zellinformation und Zell-Zellkommunikation einer umfassenden Revision bedürfen.
Was Baltimore und die große Mehrheit seiner Kollegenschaft nach dem Zusammenbruch des genetischen Weltbildes nicht sagen, ist die fundamentale Tatsache, dass alle grundlegenden Theorien der gentechnisch fixierten modernen Medizin zur Zellenergie, Zellinformation und Zell-Zellkommunikation einer umfassenden Revision bedürfen.
Der Autor dieses Beitrages hatte aufgrund der Analyse einer großen Vielfalt von evolutionsbiologischen Forschungsdaten postuliert, dass der menschliche Zellkern in Wirklichkeit als evolutionsbiologisches Erbe ein doppeltes Genom besitzt, das aus den Genkulturen zweier zellkernloser Einzeller vor Urzeiten in einem zeitgleich gebildeten Zellkern integriert, aber im Gegensatz zur damaligen Auffassung der Evolutionsforscher, nicht verschmolzen wurde. Dieses Postulat der „Zwitternatur“ der menschlichen Zellsysteme hat sich für das Verständnis von Gesundheit und Krankheit, Altern und Tod in der therapeutischen Praxis als außerordentlich fruchtbar erwiesen.
Anfang der 70er Jahre wurden aus der Tiefe des Ozeans, wo absolut kein Sonnenlicht hindringt, mit Tauchrobotern unbekannte zellkernlose Einzeller geborgen, die lange als neue Bakterienart klassifiziert wurden. Spätere umfassende Sequenzvergleiche der Nukleinsäuren und Proteine dieser Mikroorganismen ergaben jedoch fundamentale Unterschiede zu den Bakterien, sodass die Evolutionsbiologen die 5 Reiche des Lebens neu ordneten in 3 sog. Domänen: die jetzt Archaea genannten zellkernlosen Einzeller, die zellkernlosen Bacteria und die Lebewesen mit echtem Zellkern, die Eukarya (einzellige Protista, einzellige und mehrzellige Algen, einzellige und mehrzellige Pilze, Pflanzen, Tiere und der Mensch).
Entscheidend war auch die revolutionäre Erkenntnis, dass alle Eukarya, inkl. der Mensch, ihre Existenz einem einzigartigen und einmaligen Fusionsakt in der Evolutionsgeschichte verdanken, nämlich der Besiedlung einer voluminöseren Archaea-Art als Wirts- oder Stammzelle durch Einzeller aus der Domäne der Bacteria. Diese Bildung einer intrazellulären Lebensgemeinschaft aus Angehörigen der beiden unterschiedlichen Domänen und der Integration der beiden an sich nicht kompatiblen fremden Genomkulturen in einem gemeinsamen Zellkern, vom Autor als Cellsymbiosis bezeichnet, fand zu einem sehr markanten Zeitpunkt der Erdgeschichte vor 2,1 Mrd. Jahren statt. Vorausgegangen war vor 2,4 Mrd. Jahren die erste von 3 Totalvereisungen des gesamten Erdballs. Wie Geologen nachgewiesen haben, war die Erdatmosphäre vor der globalen Vereisung frei von molekularem Sauerstoff (O2), es dominierte CO2-Gas und vor allem Methangas (CH4). Das CO2 stammte aus der regen Vulkantätigkeit der Erdkruste, das Methangas (CH4) jedoch von den allgegenwärtigen Archaea, die CO2 in CH4 umsetzten. Nach Auftauen des globalen Eispanzers stieg der O2-Gehalt der Atmosphäre exponentiell an und der Methangas-Gehalt nahm exponentiell ab. Genau am zeitlichen Schnittpunkt dieser beiden atmosphärischen Gaskurven vollzog sich die Cellsymbiosis.
Die Evolutionsbiologen haben bis heute die Frage nicht beantwortet, wie sich die strikt anaeroben Archaea (wie es heute noch in den Lehrbüchern heißt), für die minimale O2-Mengen hochtoxisch sind, und ihre bakteriellen Zellsymbionten, die bereits eine O2-abhängige Atmungskette entwickelt hatten, im gleichen Milieu begegnen konnten. Das Rätsel löst sich sofort, wenn man weiß, dass sich eine bestimmte Archaea-Art unter dem allmählich zunehmenden, existenzbedrohenden O2-Gasdruck im Ozean und in der Erdatmosphäre zu fakultativen Aerobiern weiterentwickelten: Alternativ lernten diese Archaea im mäßig O2-angereicherten Milieu CH4 mit Hilfe von O2 zu verstoffwechseln und Elektronen und Protonen für die lebenswichtige Bereitstellung von Adenosintriphosphat (ATP) zu gewinnen. Dieser ATP-Stoffwechselweg ist von den Mikrobiologen in Methan-bildenden Archaea und Bakteria nachgewiesen worden. Im Sauerstoff-freien Milieu konnten dieselben Archaea jedoch überleben, indem sie die ATP-Produktion auf den allen Organismen gemeinsamen ältesten Stoffwechselweg des Glukoseabbaus (Glykolyse) umschalteten. Diese Tätigkeit der fakultativ aeroben Archaea war die entscheidende Bedingung für die Cellsymbiosis mit den bakteriellen Symbionten, die bereits eine O2-abhängige Atmungskette entwickelt hatten. Bis Ende der 90er Jahre konnten nun von den Evolutionsforschern entscheidende Befunde für die menschliche Cellsymbiosis gesichert und publiziert werden: etwa 60% der Gene im menschlichen Genom leiten sich ab von den Genen der Stammzelle der fakultativ aeroben Archaea (vom Autor als A-Genom bezeichnet). Das A-Genom ist dominant während des Zellteilungszyklus ab der S-Phase (Verdoppelungsphase des Doppelstranges der beiden DNA-Stränge zur Weitergabe je einer Doppelhelix auf eine der sich bildenden Tochterzellen). Die übrigen Gene (vom Autor als B-Genom bezeichnet) stammen (überwiegend?) von den Genen ab, die von den bakteriellen Symbionten in den gemeinsamen Zellkern delegiert wurden.
Das B-Genom ist dominant während der Phasen der differenzierten Zellleistungen, abhängig vom jeweiligen Zellleistungstyp.
Auf der Basis des hier skizzierten Szenarios konnte der Autor das Krebsgeschehen neu interpretieren. In den 20er Jahren hatte der Biochemiker und spätere Nobelpreisträger Otto Warburg erstmals das Phänomen beschrieben, dass Krebszellen trotz Anwesenheit von O2 anscheinend ihre ATP-Produktion überwiegend über Glykolyse im Zellplasmadurchführen. Dieses sog. Warburg- Phänomen wird jedoch bis heute kontrovers diskutiert, da in den Nachkommen der bakteriellen Symbionten, den zu hochkomplexen Leistungsträgern in allen Zelltypen weiterentwickelten Mitochondrien, auch in Krebszellen ein nicht unbeträchtlicher O2-Verbrauch nachgewiesen wurde. 2002 publizierten australische Krebsforscher die Ergebnisse einer präzisen Messung des tatsächlichen O2-Konsums in der üblicherweise für solche Untersuchungen eingesetzten BrustKrebszelllinie MCF-7 für die Dauer von 5 Tagen mit modernsten Sauerstoffelektroden. Gleichzeitig kritisierten die Forscher die bis dahin durchgeführten Messungen dieser Art als zu kurzfristig. Das verblüffende Resultat: Der O2-Verbrauch in diesen Krebszellen lag nicht wesentlich niedriger als in vielen intakten differenzierten Zelltypen, die Glykolyse nicht wesentlich höher. Aber: Die Forscher konnten 65% der Stoffwechselsubstrate zur Gewinnung der Elektronen und Protonen für die O2-abhängige ATP-Produktion nicht identifizieren. (Guppy M. et al. Contribution to different fuels and metabolic pathways to the total ATP turnover of poliferating MCF-7 breast cancer cells. Biochem J. (2002), May 15; 364 (Pt 1): 309-15)
Diese Befunde demonstrieren, dass die „Zwitternatur” der menschlichen Zellsysteme bis heute von den klinischen Krebsforschern überhaupt nicht verstanden worden ist. Um dieses Dilemma zu lösen, hat der Autor die begründete Annahme eingeführt, dass das Krebsgeschehen sozusagen wie in einem Rückspiegel die Entwicklungsphasen der Evolution widerspiegelt: die funktionelle Störung der Regulationsebene der aeroben O2-Nutzung zur ATP-Produktion über das enzymatische Oxidase-System in den Mitochondrien erzwingt eine protektive Umschaltung auf die Regulationsebene der fakultativ aeroben O2-Nutzung zur ATP-Produktion über das enzymatische Oxygenase-System im Zellplasma. Eine solche evolutionsbiologisch programmierte Schutzschaltung kann erstmalig den bisher nicht identifizierten Substratanteil für die O2-abhängige Bereitstellung von Elektronen und Protonen in den Tumorzellkolonien und auch das Warburg-Phänomen erklären.
 Warburg hatte ein Entweder-Oder postuliert, da er einen strukturellen Defekt im Cytochromoxidase-Komplex der Atmungskette der Mitochondrien angenommen hatte: Entweder O2-Atmung in den intakten, differenzierten Zellen in den Mitochondrien, oder Glykolyse ohne O2-Nutzung trotz Anwesenheit von O2 im Zellplasma. Dem Postulat des doppelten Genoms muss jedoch modellgemäß das Postulat des doppelten O2-Nutzungssystems zugeordnet werden. Unter überdauerndem chronischem Zellstress vielfältiger Natur können sich teilungsaktive Zellen in das evolutionsbiologisch ältere Zwischenstadium der ATP-Produktion zurückbilden: sowohl ATP-Produktion mit O2-Nutzung in den Mitochondrien und im Zellplasma mit unterschiedlichen Anteilen als auch ATP-Produktion durch Glykolyse ohne O2-Nutzung im Zellplasma, letztere anteilsmäßig abhängig vom Regressionszustand der sich bildenden Krebszelle. Das B-Genom verliert schrittweise die Kontrolle über die differenzierten Zellleistungen zuungunsten einer zunehmenden Dominanz des A-Genoms als archaisch programmierte Überlebensstrategie.
Warburg hatte ein Entweder-Oder postuliert, da er einen strukturellen Defekt im Cytochromoxidase-Komplex der Atmungskette der Mitochondrien angenommen hatte: Entweder O2-Atmung in den intakten, differenzierten Zellen in den Mitochondrien, oder Glykolyse ohne O2-Nutzung trotz Anwesenheit von O2 im Zellplasma. Dem Postulat des doppelten Genoms muss jedoch modellgemäß das Postulat des doppelten O2-Nutzungssystems zugeordnet werden. Unter überdauerndem chronischem Zellstress vielfältiger Natur können sich teilungsaktive Zellen in das evolutionsbiologisch ältere Zwischenstadium der ATP-Produktion zurückbilden: sowohl ATP-Produktion mit O2-Nutzung in den Mitochondrien und im Zellplasma mit unterschiedlichen Anteilen als auch ATP-Produktion durch Glykolyse ohne O2-Nutzung im Zellplasma, letztere anteilsmäßig abhängig vom Regressionszustand der sich bildenden Krebszelle. Das B-Genom verliert schrittweise die Kontrolle über die differenzierten Zellleistungen zuungunsten einer zunehmenden Dominanz des A-Genoms als archaisch programmierte Überlebensstrategie.
In diesem Zusammenhang kann auch erklärt werden, warum seit Ausrufung des „Krieges gegen den XXX (Nachfolgernder Abschnitt muß aus Gründen HWG § 12 A2 gestrichen werden…)
Dagegen hat die vom Konzept der Cellsymbiosis abgeleitete Therapieform bei vielen chronischen Erkrankungen beeindruckende Behandlungserfolge gezeitigt (Lowenfels, D. (2006). The Dual Strategy of the Immune Response. A Review of Heinrich Kremer’s Research on the Pathophysiology of xxx and Other Chronic Immune Imbalances. Townsend Letter. Dies gilt nicht nur für sog. austherapierte Patienten, sondern auch für zelluläre und humorale Immunschwächen, inflammatorische Erkrankungen, Autoimmunerkrankungen, Herzerkrankungen, Arteriosklerose, Diabetes auch in der therapieresistenten Form, Osteoporose, Burn out-Syndrom, CFS, Fibromyalgie, neurodegenerative Erkrankungen inkl. Alzheimer’sche Krankheit und andere Demenzformen sowie Parkinson’sche Krankheit, Depressionen, Psychosen, und viele andere, primär als Mitochondriopathien einzustufende Symptomzustände und Leistungsschwächen. Zielführend für die Entwicklung der Rezepturen der Cellsymbiosistherapie war die Erkenntnis des Autors, dass kurz gesagt, entgegen den bisher gültigen Theorien, die Atmungsketten in den Mitochondrien als Lichtquanten-(Photonen) Prozessoren arbeiten. (Kremer, H. Das Geheimnis-Kurzschluss im Photonenschalter 2004
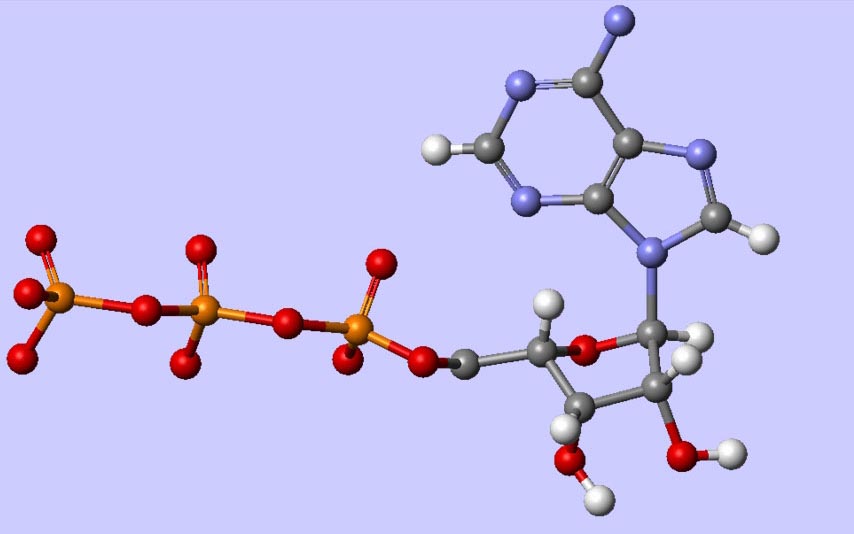
Die dabei generierte mehrdimensional modulierte Information wird auf die delokalisierten Elektronen der Doppelbindungen des Adeninmoleküls des Adenosintriphosphats transferiert. So erklärt sich, warum ATP praktisch bei allen Stoffwechselvorgängen direkt oder indirekt „aktivierend“ oder besser “informierend” beteiligt sein muss. So müssen beispielsweise die komplex modulierten Nukleobasen der ATP vor jeder Neusynthese einer DNA- oder RNASequenz die benötigten Nukleobasen-Bausteine „informieren“, d.h. codierte Schwingungsenergie auf diese übertragen.
Die eingangs zitierte hilfesuchende Frage des Genetikers Baltimore “was uns unsere Komplexität verleiht”, beantwortet sich im Prinzip so: Information ist eine nichtmaterielle Größe, die von einer Raum-Zeit-unabhängigen Matrix potentieller Information an unsere „Antennenmoleküle“ wie ATP über quantendynamische Vorgänge als „kreative Information“ übermittelt wird.
Zellen sind also keine bloßen Wärmekraftmaschinen, sondern informationswandelnde Medien. Aber ATP ist nicht gleich ATP, in den menschlichen Mitochondrien modulierte ATP-Information ist sicher komplexer moduliert als beispielsweise in der Maus. Doch unter fakultativ aeroben Bedingungen modulierte ATP ist sicher weniger komplex moduliert als mitochondriale ATP aus intakten differenzierten Zellen, oder unter glykolytischen anaeroben Bedingungen modulierte ATP ist sicher am wenigsten komplex moduliert. Im letzteren Fall sprechen auch die Krebsforscher folgerichtig von “entdifferenzierten” Zellen.
Der Genetiker Baltimore sollte sich fragen, warum nach der Abschrift einer protein-codierenden DNA-Sequenz in eine mobile Boten-RNA-Sequenz, nach Bearbeitung derselben, an die „reife“ Boten-RNA ein sog. Poly-A-Schwanz angehängt werden muss, weil sonst die Proteinsynthese nicht funktioniert. Die Anweisung dazu ist nicht in den Genen zu finden.
Woher weiß die Zelle also, was sie zu tun hat? Die Antwort an Baltimore lautet: Weil die ca. 270 Adeninmoleküle des Poly-A-Schwanzes, die von modulierter ATP abstammen, resonanzgekoppelt sind mit dem nichtmateriellen Informationsfeld. Stellt man sich diesen Poly-A-Schwanz also als unterschiedlich Lichtquanten-modulierte Adenin- Elemente vor, dann ergibt sich ein codiertes Lichtquantenmuster, und so kann man sich den gesamten Organismus als hochkomplexes „informiertes“ Lichtquantenfeld vorstellen (als quantendynamische Modellvorstellung siehe Bohm. D (1990)
A new theory of the realtionship of mind and matter. Philosophical Psychology: Vol. 3 N. 2.271-86)
In der Cellsymbiosistherapie werden deshalb Naturstoffe eingesetzt, die über bestimmte Wellenlängen bzw. Frequenzfenster im nahen UV-Bereich und im sichtbaren Spektrum Lichtquanten absorbieren und emittieren. Das therapeutische Potential solcher Naturstoffe ist in neueren Forschungspublikationen bestätigt worden.
(Middlestone, E., Jr, et al (2000) The effekts of plant flavonoids on mammalian cells: implications for Inflammation, heart disease, and XXXXXX. Pharmacol. Res. 52,673-751; Aggarwal B.B. et al. (2003) AntiXXXXXX potential of curcumin: preclinical and clinical studies.
Insbesondere wird das Konzept der Cellsymbiosistherapie gestützt durch die neueren faszinierenden Ergebnisse der experimentellen und klinischen Grundlagenforschung über Alterungsprozesse. Im Zusammenhang mit der Neuentdeckung der Enzym-Klasse der sog. Sirtuine (von engl. Silent information regulator), die bestimmte Gene und Proteine durch Entfernen einer aktivierenden Molekülgruppe stummschalten, wurden verblüffende Effekte bei allen Eukarya erkannt. So wurden beispielsweise die Sirtuin-Enzyme von für xxxxx und Diabetes besonders disponierten Mäusen aktiviert mit bestimmten Naturstoffen aus der großen Familie der pflanzlichen Polyphenole. Im Vergleich zu normalen Kontrollmäusen lebten die disponierten Mäuse wesentlich länger und entwickelten trotzdem selten xxxxx, Diabetes oder neurodegenerative Erkrankungen.
Diese Forschungsdaten belegen, dass es auch beim Menschen ein übergeordnetes Regulationssystem gibt, da Sirtuin-Enzyme inzwischen auch im Zellkern, im Zellplasma und in den Mitochondrien des Menschen nachgewiesen wurden. Im Ergebnis aktivieren die Lichtquanten-absorbierenden pflanzlichen Polyphenole über vielfach vernetzte Regelkreise die O2-abhängige Mitochondrienleistung. Das lange gehegte wissenschaftliche Vorurteil, die Alterungsprozesse und die damit verbundenen typischen Krankheiten, wie Krebs, Diabetes, Herzkreislauferkrankungen und neurodegenerative Krankheitsformen, seien ein unvermeidbarer natürlicher Verschleißprozess, ist damit in Frage gestellt.
(Wood, J.G. et al. (2004) Sirtuin activators mimic caloric restriction and delay aging in metazoans. Nature 430, 686-89; Porcu, M., Chiarugi, A. Sirtuin-interacting drugs: from cell death to lifespan extension. Trends in Pharmacological Sciences, Vol. 26 N. 2 February 2005; Sinclair, D.A., Guarente, L., Schlüssel zur Langlebigkeit. Spektrum der Wissenschaft. Oktober 2006)
Strukturanaloge Lichtquanten-modulierende pflanzliche Polyphenole, garantiert rückstandsfrei und unbelastet von Schwermetallen und Schadstofffen, sind in kombinierter und besonderer galenischer Zubereitungsform ein Hauptbestandteil der Rezepturen der Cellsymbiosistherapie. Polyphenole können vom Säugetierorganismus nicht synthetisiert werden und besitzen deshalb für den Menschen Vitamincharakter. Sie sind für die intakte Mitochondrienfunktion essentiell. Aus diesem Grunde sind pflanzliche Polyphenole in geeigneter Kombination mit anderen Naturprodukten zur Verhütung und Behandlung schwerwiegender mitochondrialer Leistungsschwächen, Systemerkrankungen und vorzeitiger Alterungsprozesse indiziert. Sie werden in einem individuellen Vorbeugungs- oder Behandlungskonzept als sog. Nahrungsergänzungsmittel von Ärzten und Heilpraktikern therapeutisch verordnet.
Copyright 2007 by Dr. med. Heinrich Kremer, Barcelona – Erstveröffentlichung in: OM und Ernährung, Juli 2007
Falls Sie eine Übersicht über die neuen vom Arzt und Forscher, Dr. Heinrich Kremer erforschten und verwendeten Mittel haben möchten, schreiben Sie mir einfach kurz eine E-Mail.




Info zum Seminar:
„Die grundlegende Bedeutung des lichtquanteninformatorischen Modells im Konzept der Cellsymbiosistherapie“
Vor 110 Jahren entdeckte der Physiker und spätere Nobelpreisträger Max Planck, dass elektromagnetische Strahlung nicht kontinuierlich, sondern in winzigsten Energieportionen oder „Quanten“ abgestrahlt wird. In den zwanziger Jahren des vorigen Jahrhunderts entwickelten u.a. die Physiker Niels Bohr, Werner Heisenberg, Paul Dirac und Erwin Schrödinger, die ebenfalls mit dem Nobelpreis ausgezeichnet wurden, die „Quantenmechanik“. Primär ging es um die mathematische Modellierung der bis dahin völlig rätselhaften Naturkräfte, welche die Interaktionen zwischen den Protonen und Neutronen in den Atomkernen der Elemente und den sie auf komplizierten Bahnen umkreisenden Elektronen bestimmen.
Zum damaligen Zeitpunkt konnten die bizarren theoretischen Annahmen und dem Alltagsverstand zuwiderlaufenden Voraussagen der Quantenphysik im Experiment noch nicht auf ihre Gültigkeit überprüft werden. Im Laufe der folgenden Jahrzehnte sollte sich jedoch in zahllosen ausgeklügelten Experimenten erweisen, dass die Quantenfeldtheorie die am präzisesten bestätigte Theorie der modernen Physik überhaupt ist.
Bereits 1935 hatte der wohl berühmteste Physiker des 20. Jahrhunderts, Albert Einstein, zusammen mit seinen Physikerkollegen Podolsky und Rosen, in einem Fachaufsatz die fundamentale Frage aufgeworfen, ob die Quantenphysik als „vollständig“ angesehen werden könne. In einem berühmt gewordenen Gedankenexperiment hatten sie aus dem Algorithmus, also der Rechenvorschrift der Quantenmechanik, die Konsequenz abgeleitet, dass unter bestimmten Bedingungen quantisierte Elementarteilchen sich „nicht-lokal“ gegenseitig beeinflussen müssten. Das bedeutete, dass der Kausalbegriff der klassischen Physik aufgehoben wäre. Dieser besagt, dass die Einwirkung einer Kraft als Ursache auf die Veränderung eines gegebenen physikochemischen Zustandes als Wirkung nur dann vonstatten gehen kann, wenn in einer Ursache-Wirkungskette ein raum-zeitlich beschreibbarer physischer Kontakt gegeben ist.
Für Einstein und alle Physiker der damaligen Zeit, einschließlich der Väter der Quantenphysik, war eine „nicht-lokale Realität“, die Einstein als „spukhafte Fernwirkung“ verspottete, in der vierdimensionalen Raumzeit des Universums undenkbar. Aus dieser unanfechtbaren Position der Grundüberzeugung der Physikergemeinde zogen Einstein und seine Kollegen den Schluss, dass die Quantenmechanik unvollständig sein müsse und in den Voraussagen des Algorithmus ein verborgener Fehler stecken müsse. Der damalige „Papst“ der Quantenphysik, der Däne Niels Bohr, antwortete Einstein postwendend. Auf den eigentlichen Kern des naturwissenschaftlichen Problems ging er nicht näher ein. Er bestätigte die ausschließliche Existenz der „lokalen Realität“ und philosophierte eher weitschweifig über Subjekt und Objekt in der Quantenphysik. Physiker lästerten denn auch alsbald über den „großen Nebel aus dem hohen Norden“. Dieser nebulöse Zustand der Quantenfeldtheorie lichtete sich erst fast drei Jahrzehnte später, als der irische Physiker John Bell, der damals am Europäischen Kernforschungszentrum in Genf forschte, die mathematisch stringente Hypothese untersuchte, ob es in der klassischen Physik einen Algorithmus geben könne, der die gleichen experimentell überprüfbaren Voraussagen ermöglichen würde, wie der Algorithmus der Quantenmechanik. Wäre dies der Fall, dann hätten sich die Quantenphysiker geirrt und der geniale Einstein hätte die ausschließlich lokale Realität allen Seins in unserem Universum richtig diagnostiziert.
Doch es kam anders. Bell führte den mathematischen Beweis, dass es keinen klassischen Algorithmus geben kann, der die gleichen Voraussagen macht, wie die Quantenmechanik. 1981 gelang erstmalig französischen Physikern in einem quantenoptischen Experiment nach Ausschluss der sog. Bell’schen Ungleichungen der exakte Nachweis der Existenz der nicht-lokalen Realität. In der Folge haben quantenphysikalische Forschungsgruppen aus mehreren Ländern in immer ausgeklügelteren Experimenten beispielsweise mit sog. verschränkten Photonenpaaren den Nachweis der nicht-lokalen, im Sinne der klassischen Physik akausalen, Wirklichkeit bestätigen können.
Trennten die Forscher solche Lichtquantenpaare und führten eine Messung an einem der Lichtquanten durch, beispielsweise durch Bestimmung der Orientierung der Schwingungsebene des Lichtquantenzustandes (Polarisation), so stellte sich die Polarisation des beliebig entfernten anderen Photons ohne jeden Zeitverzug (instantan) entsprechend ein, völlig unabhängig vom vorhergehenden Zustand des Quantenparameters. Im Klartext: die Photonen existieren als komplex überlagerte Schwingungszustände im Quantenfeld der elektromagnetischen Strahlung ohne festgelegte Eigenschaften (virtuell).
Das bedeutet: die Photonen repräsentieren die noch ungeprägte Quanteninformation, die erst durch Interaktion mit der materiellen Matrix, die nach der Vorstellung der Quantenphysik nichts anderes ist als „gefrorene“ oder „geronnene“ Quantenzustände, ihre individuellen, real messbaren Eigenschaften gewinnt. Der Unterschied zwischen den materiellen Elementarteilchen wie beispielsweise Protonen und Elektronen sowie den Photonen besteht darin, dass erstere Masse und Ladung besitzen und letztere weder Ruhemasse noch Ladung aufweisen, aber als Austauschkräfte im Standardmodell der Elementarteilchenphysik der strukturkonservativen „okalen Realität“ fortwährend unerschöpflich kreative Quanteninformation aus der raumzeitlich dimensionslosen, nicht-lokalen Wirklichkeit, eben durch Einsteins „spukhafte Fernwirkungen“, aufmodulieren können. Diese erkannte und experimentell bewiesene Dynamik im Makrokosmos wie im Mikrokosmos ist der Grund, warum die Quantenforscher davon sprechen, dass Information die Wirklichkeit bestimmt und nicht, wie die meisten Naturwissenschaftler und Mediziner immer noch glauben, Information ein zufälliges Beiprodukt von Energie und Materie ist.
Im Spezialseminar der Cellsymbiosis werden die Grundbegriffe und Grundlagenerkenntnisse der informatorischen Quantenwelt genauer erklärt und veranschaulicht. Es werden sodann im präzisen Detail die bisher nicht erklärten Strukturen und Funktionen analysiert, die innerhalb und zwischen den menschlichen Zellsystemen und systemisch übergreifend das Prinzip der Cellsymbiosis ermöglichen, nämlich in jedem Augenblick die psychische und physische Integrität zu bewahren oder wiederherzustellen durch Auslese, Speicherung, Modulation, Transfer, Verarbeitung und Resonanzkopplung von Quanteninformation aus der nicht-lokalen Wirklichkeit. Dabei soll deutlich werden, dass es zunächst einmal zu verstehen gilt, was Gesundheit bedeutet, um den „Störfall“ Krankheit nachvollziehen zu können.
Folgendes auszugsweises Zitat möge veranschaulichen, wie groß zur Zeit die Verwirrung und Ratlosigkeit beispielsweise in der genfixierten pharmamedizinischen Forschung ist, weil man völlig die Tatsache verdrängt hat, dass der Geist sich über Quanteninformation vermittelt. Dies setzt zwar die intakte biologische Matrix voraus, die aber ständig der Identität stiftenden Leitinformation bedarf.
„Die Vorstellung, man müsse nur genügend DNA-Sequenzen buchstabieren, um ans Ziel zu kommen, wird mit jeder neuen Genstudie ohnehin immer fragwürdiger. Was in einer lebenden Zelle geschieht, lässt sich nicht durch eine lineare Abfolge von (molekularen) Buchstaben erklären. Die Biologie macht damit ähnliche Erfahrungen wie vor ihr schon die Physik: Elementarteilchen lassen sich finden, wenn man nur genau genug hinsieht. Sie nützen nur nichts für die Wettervorhersage“. (Jörg Albrecht: Und nun zum Wetter. Frankfurter Allgemeine Sonntagszeitung, 7. 9. 2008, Nr. 36, S.65).
Mit der abschließenden sarkastischen Bemerkung will der Autor sagen, dass auch das Wetter den Gesetzen einer nicht-linearen Dynamik gehorcht.
Mir anderen Worten: Die moderne Medizin hat kein schlüssiges und plausibles Grundlagenkonzept. Je weniger die Medizinforscher von der menschlichen Cellsymbiosis verstanden haben, haben sie sich in die „babylonische Gefangenschaft“ der Pharmaindustrie begeben.
Ein abschließendes Zitat: „Im Rückblick waren unsere damaligen Annahmen über die Funktionsweise des Genoms dermaßen naiv, dass es fast schon peinlich ist.“ (zitiert in Ulrich Bahnsen: Erbgut in Auflösung. Das Genom galt als unveränderlicher Bauplan des Menschen, der zu Beginn unseres Lebens festgelegt wird. Von dieser Idee muss sich die Wissenschaft verabschieden.
In Wirklichkeit sind unsere Erbanlagen in ständigem Wandel begriffen. Zeit Online 25/2008 S.33). Des Zitat stammt von dem US-Unternehmer Craig Venter, der bei der computergestützten Sequenzierung des vollständigen menschlichen Genoms im Wettrennen mit tausenden staatlichen Genforschem aus 20 Nationen des Humanen Genomprojektes 2001 als Erster die vorläufigen Ergebnisse vorlegen konnte. Das Time Magazine schrieb damals: „Dank dem aggressiven und abgebrühten Craig Venter kann die Welt die Partitur des menschlichen Genoms lesen“ (Werbung zur deutschen Ausgabe des Buchs von Craig Venter: Entschlüsselt. Mein Genom, mein Leben. November 2009). In Wirklichkeit sind die Mythen der Genomforschung längst dahingeschmolzen wie der Schnee von gestern.
Fachliche Fräsen und Anregungen zu diesem Spezialseminar sollten frühzeitig vor Seminarbeginn über die Akademie Cellsymbiosistherapie eingebracht werden, um sie den übrigen Teilnehmern und dem Referenten rechtzeitig zur Kenntnis zu geben.
Dr. med. Heinrich Kremer, Barcelona, November 2009
Studienlage:
Studienrechercheergebnisse bzgl. der diagnostischen/therapeutischen Aspekte der Cellsymbiosistherapie® – Konzeptes nach Dr. med. Heinrich Kremer – die Atmungskette- und Mitochondrien-Therapie
(Quelle: www.pubmed.gov – weltweit größte Fachbibliothek bzgl. Medizin und Forschung)
Stand 2007
I. Schwermetalle
| Anzahl der Studien | |
| Einfluss von Schwermetallen auf die Atmungskette | 1.826 |
| Einfluss von Quecksilber auf die Atmungskette | 62 |
| Einfluss von Schwermetallen bzgl. Krebs und die Atmungskette | 19 |
| Einfluss von Schwermetallen auf die Mitochondrien | 4.495 |
| Schwermetalle und Krebs | 23.762 |
| Einfluss von Quecksilber auf die Mitochondrien | 296 |
II. Bedeutung von verschiedenen Polyphenolen für die Mitochondrien/Atmungskette
| Anzahl der Studien | |
| Oligomere Procyanidine (OPC) | 32 |
| Quercetin und Mitochondrien | 94 |
| Quercetin und Atmungskette | 19 |
| Quercetin | 5.449 |
| Curcumin und Mitochondrien | 75 |
| Curcumin und Atmungskette | 1 |
| Curcumin | 2.023 |
| Agaricus blazii muril und Mitochondrien | 2 |
| Agaricus blazii muril | 47 |
| Agaricus blazii muril und NO-Gas | 1 |
III. Bedeutung der Supra-Spurenelemente für die Mitochondrien/Atmungskette
| Anzahl der Studien | |
| Chrom und Atmungskette | 11 |
| Cytochrom und 4. Komplex der Atmungskette | 27 |
| Chrom und Mitochondrien | 45 |
| Vanadium und Mitochondrien | 64 |
| Vanadium und Atmungskette | 23 |
| Vanadium | 4.334 |
| Molybdän und Mitochondrien | 50 |
| Molybdän und Atmungskette | 169 |
| Molybdän | 6.228 |
| Molybdän und Krebs | 277 |
| Mangan und Mitochondrien | 1.092 |
| Mangan und Atmungskette | 300 |
| Mangan | 25.389 |
| Mangan und Krebs | 1.329 |
| Chrom | 205 |
| Chrom und Krebs | 6 |
| Vanadium und Krebs | 196 |
IV. Bedeutung der Aminosäuren für die Mitochondrien/Atmungskette
| Anzahl der Studien | |
| BCAAs (L-Leucin, L-Isoleucin und L-Valin) und Mitochondrien | 1.075 |
| BCAAs (L-Leucin, L-Isoleucin und L-Valin) und Atmungskette | 89 |
| BCAAs (L-Leucin, L-Isoleucin und L-Valin) | 38.678 |
| BCAAs (L-Leucin, L-Isoleucin und L-Valin) und Krebs | 2.717 |
| Glutathion und Mitochondrien | 3.251 |
| Glutathion und Atmungskette | 460 |
| Glutathion | 77.029 |
| Glutathion und Krebs | 10.268 |
| Methionin und Mitochondrien | 1.030 |
| Methionin und Atmungskette | 175 |
| Methionin | 57.274 |
| Methionin und Krebs | 5.315 |
| Cystein und Mitochondrien | 2.152 |
| Cystein und Atmungskette | 376 |
| Cystein | 79.567 |
| Cystein und Krebs | 6.787 |
| Arginin und Mitochondrien | 914 |
| Arginin und Atmungskette | 210 |
| Arginin | 81.631 |
| Arginin und Krebs | 3.636 |
V. Bedeutung weiterer Substanzklassen für die Mitochondrien/Atmungskette
| Anzahl der Studien | |
| Folsäure und Mitochondrien | 186 |
| Folsäure und Atmungskette | 22 |
| Folsäure | 28.799 |
| Folsäure und Krebs | 8.057 |
| Fettsäuren und Mitochondrien | 8.285 |
| Fettsäuren und Atmungskette | 963 |
| Fettsäuren | 297.594 |
| Fettsäuren und Krebs | 16.808 |
| Phospholipide und Mitochondrien | 5.048 |
| Phospholipide und Atmungskette | 623 |
| Phospholipide | 127.451 |
| Phospholipide und Krebs | 7.127 |
| Cardiolipin und Mitochondrien | 954 |
| Cardiolipin und Atmungskette | 110 |
| Cardiolipin | 5.441 |
| Cardiolipin und Krebs | 181 |
| Carnitin und Mitochondrien | 2.410 |
| Carnitin und Atmungskette | 183 |
| Carnitin | 9.643 |
| Carnitin und Krebs | 218 |
| Ubichinon (Q 10) und Mitochondrien | 1 |
| Ubichinon (Q 10) | 3 |
VI. Lebensmittelantikörpertest / Leaky Gut / Glutenunverträglichkeit
| Anzahl der Studien | |
| IGG4 | 98.939 |
| IGG4 und Krebs | 9.689 |
| IGG4 und Histamin | 729 |
| IGG4,Krebs und Histamin | 25 |
| Lebensmittelallergie („food allergy“) | 10.647 |
| Lebensmittelallergie („food allergy and intolerance“) | 945 |
| Lebensmittelunverträglichkeit („food intolerance“) | 3.09 |
| Leaky Gut | 36 |
| Leaky Gut und Zitronensäure | 35 |
| Leaky Gut und IGG4 | 1 |
| Leaky Gut und Schwermetalle | 1 |
| Glutenintoleranz | 11.43 |
| Glutenintoleranz und Krebs | 963 |
| Gluten und Aspergillus Niger (AN) | 6 |
| Gluten und Krebs | 288 |
| Aspergillus Niger (AN) | 4.263 |
| AN und Krebs | 49 |
| AN und Immunsystem | 35 |
| AN und Mitochondrien | 42 |
| AN und Atmungskette | 21 |
| AN und Schwermetalle | 205 |
| AN und Dünndarm | 10 |
| AN und Dickdarm | 5 |
| AN und Cardiolipin | 2 |
| AN und Arginim | 20 |
| AN und Formaldehyd | 12 |
| AN und Glutathion | 24 |
| AN und BCAA | 12 |
| AN und Zyanid | 29 |
| AN und Methan | 9 |
| AN und CD4 Zellen | 3 |
| Pilze und Krebs („fungus and cancer“) | 6.343 |
| AN und Obduktion von Krebsverstorbenen | 3 |
| AN und Obduktion | 8 |
VII. NO-Gas (Stickoxid-Gas)
| Anzahl der Studien | |
| NO-Gas | 83.557 |
| NO-Gas undund Arginin | 24.590 |
| Arginin | 81.709 |
| NO-Gas und Cystein | 1.949 |
| Cystein | 79.652 |
| NO-Gas und Glutamin | 252 |
| Glutamin | 27.460 |
| NO-Gas und Glycin | 387 |
| Glycin | 51.779 |
| NO-Gas und Glutathion | 2.922 |
| Glutathion | 77.130 |
| NO-Gas und Mitochondrien | 1.665 |
| NO-Gas und Atmungskette | 495 |
| NO-Gas und Krebs | 3.526 |
| NO-Gas und Schwermetalle | 1.738 |
| NO-Gas und Quecksilber | 80 |
| NO-Gas und Arsen | 66 |
| Arsen | 12.068 |
| NO-Gas und Fettsäuren | 6.285 |
| NO-Gas und Phospholipide | 837 |
| NO-Gas und Cardiolipin | 16 |
| Phospolipide | 127.542 |
| NO-Gas und BCAA | 247 |
| NO-Gas und Hämoxygenase | 683 |
| NO-Gas und VirusInfektionen | 851 |
| NO-Gas und Darm | 2.643 |
| NO-Gas und Methan | 65 |
| NO-Gas und Zyanid | 4.306 |
| NO-Gas und Sirtuine | 3 |
| NO-Gas und Quercetin | 174 |
| NO-Gas und Ellaginsäure | 9 |
| NO-Gas und Folsäure | 123 |
| NO-Gas und Mangan | 4 |
| NO-Gas und Molybdän | 39 |
| NO-Gas und Sekret. IGA | 7 |
| Sekret. IGA | 47 |
| Sekret. IGA und Schwermetalle | 25 |
| Sekret. IGA und AN | 1 |
VIII. Weiteres
| Anzahl der Studien | |
| Stammzellen | 150.550 |
| Stammzellen und Krebs | 31.849 |
| Tumorstamzellen | 4.231 |
| Krebs und Glutathion | 10.268 |
| Stammzellen und Sirtuine | 19 |
| Sirtuine | 638 |
| Sirtuine und NO-Gas | 3 |
| Sirtuine und Glutathion | 8 |
| Sirtuine und Arginin | 7 |
| Sirtuine und Cystein | 4 |
| Sirtuine und BCAA | 1 |
| Sirtuine und Interleukine | 2 |
| Interleukine | 161.561 |
| Sirtuine und CD4 Zellen | 1 |
| Sirtuine und Immunsystem | 8 |
| Sirtuine und Sauerstoff | 33 |
| Sirtuine und Nervenzellen | 21 |
| Sirtuine und ATP | 8 |
| Sirtuine und Mitochondrien | 32 |
| Effektive Mikrooganismen (EM) | 438 |
| EM und Darm | 24 |
| EM und IGG4 | 10 |
| EM und Sekret. IGA | 3 |
| EM und Dünndarm | 2 |
| Laktobacillus | 14.823 |
| Laktobacillus und Dünndarm | 412 |
| Laktobacillus und Dickdarm | 529 |
| Fruktooligosaccharide | 205 |
| Fruktooligosaccharide und Dünndarm | 24 |
| Fruktooligosaccharide und Dickdarm | 55 |
| Alkalose und Krebszellen | 78 |
| MCF 7 Krebszellen und Sauerstoff | 172 |
| Molybdändefizit und Krebs | 30 |
| Mangandefizit und Krebs | 51 |
| Chrom und Krebs | 277 |
| Mangan und Krebs | 1.329 |
| Cytotoxische Effektorzellen | 7.608 |
| Cytotoxische Effektortellen und Krebs | 2.745 |
| Cytotoxische Effektorzellen und Schwermetalle | 62 |
| CD 4 Zellen | 17.072 |
| CD 4 Zellen und Krebs | 4.139 |
| CD 4 Zellen und Schwermetalle | 795 |
| Hämoxygenase | 2.484 |
| Hämoxygenase und Schwermetalle | 409 |
| Hämoxygenase und Krebs | 88 |
| Hämoxygenase und Methan | 2 |
| Hämoxygenase und Zyanid | 58 |
Studien Gesamt: 2.126.715



